
There is a gap in the fossil record just before the point where we’d expect to find our earliest ancestors, a window between roughly 10 and 6 million years ago when the lineage leading to humans was presumably doing something interesting locomotively. What we find in that window is mostly silence, punctuated by fragmentary teeth and jaw pieces from Africa and, increasingly, from unexpected places in Europe and western Asia.
The Azmaka site in southern Bulgaria, near the town of Chirpan, sits in the Upper Thracian Plain. It has been producing fossils for decades, mostly from the Late Miocene, an extinct world of three-toed horses, early mastodons, and an extraordinarily rich savannah fauna. In 2016, excavators working the AZM-6 sediment horizon pulled out a nearly complete right femur. A thighbone. They didn’t know what it was at first, in the sense that its taxonomic identity required careful argument. But its anatomy was remarkable from the start.
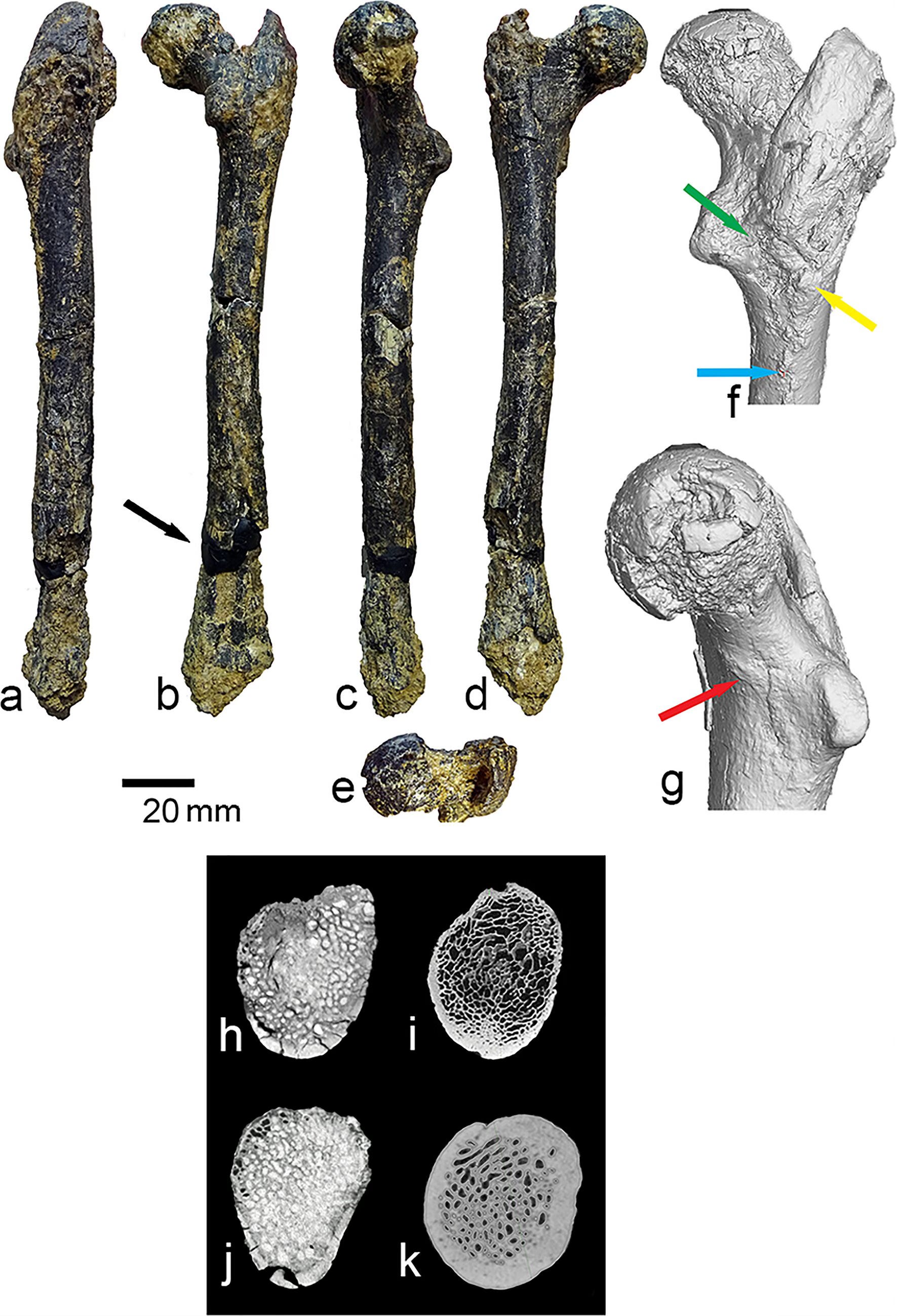
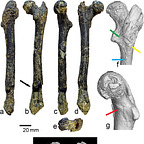
That femur, catalogued as FM3549AZM6, is now the subject of a detailed new analysis published in Palaeobiodiversity and Palaeoenvironments1 by a team led by Nikolai Spassov of Bulgaria’s National Museum of Natural History, along with Dionisios Youlatos, Madelaine Böhme, and David Begun. Their argument, stated plainly: this bone belongs to a hominine that was walking bipedally, at least part of the time, 7.2 million years ago, in what is now the Balkans.
The date alone should give you pause. Orrorin tugenensis, found in Kenya, has long been considered the oldest convincing biped, at about 6 million years. Sahelanthropus tchadensis, from Chad, is older at around 7 million years, but its status as a biped has recently come under serious scrutiny. A 2025 analysis by Cazenave and colleagues argued there are no definitive indicators of bipedalism in the Sahelanthropus femur. If they are right, and if the Azmaka team is right about their Bulgarian bone, then the oldest known biped walked not in Africa but in southeastern Europe.
What the bone actually looks like
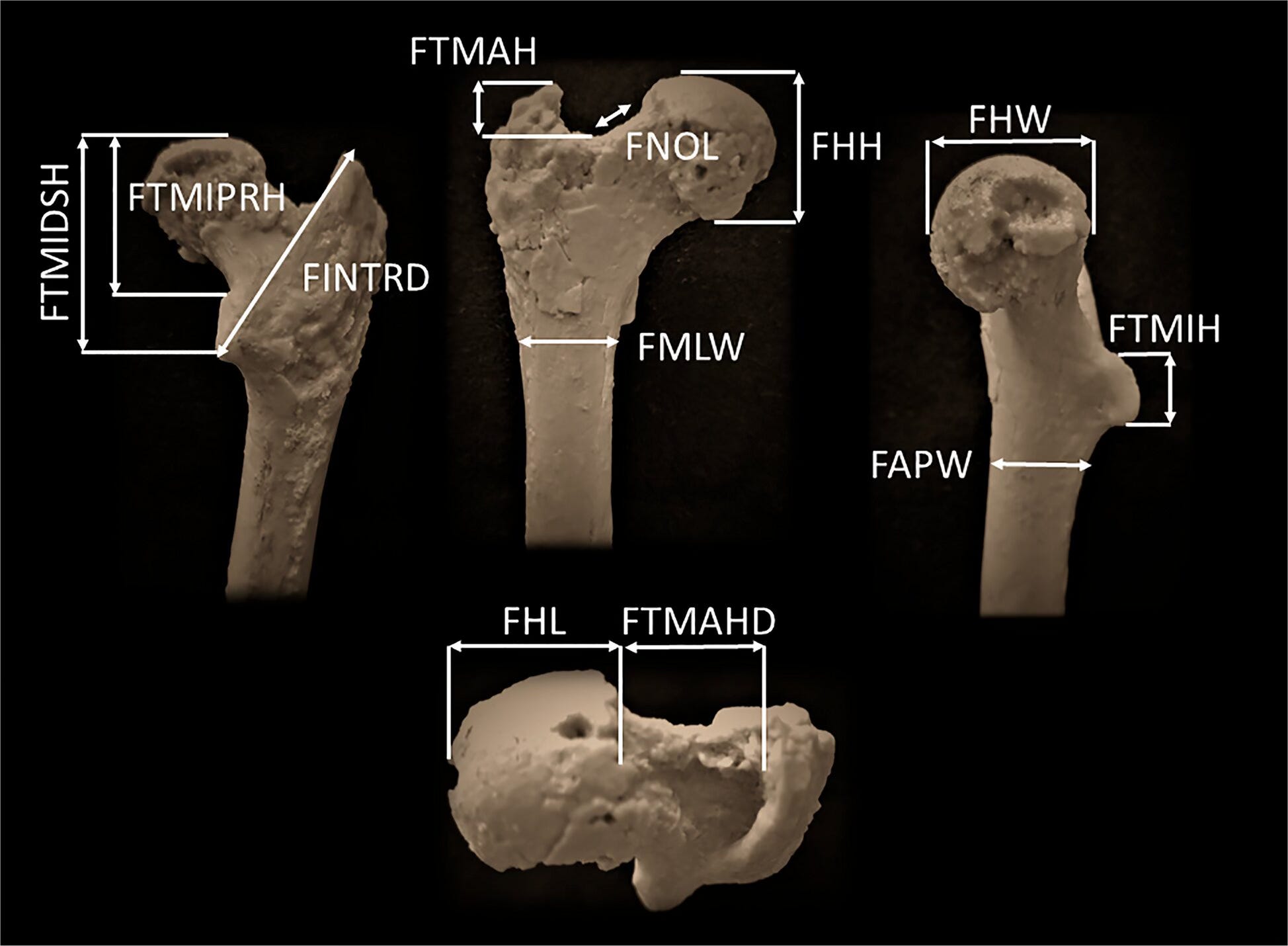
The femur is mostly complete. The distal epiphysis is missing, and there’s some diagenetic damage, but enough survives to take reliable measurements across fifteen variables of the proximal end, and enough shaft to characterize its overall shape. The bone was scanned at the Max Planck Institute for Evolutionary Anthropology in Leipzig. The team ran principal component analyses, canonical variates analyses, discriminant function analyses, and compared the fossil against an extensive reference collection spanning gibbons, orangutans, gorillas, chimpanzees, bonobos, cercopithecoid monkeys, and a range of fossil hominoids from the Miocene.
Body mass, estimated from femoral head dimensions using regressions calibrated on chimpanzees, comes in around 23 to 24 kilograms. That’s small. It’s consistent with a female individual. For comparison, the holotype mandible of Graecopithecus freybergi from Pyrgos Vasilissis in Greece is estimated to belong to a male closer to 40 kilograms. The team attributes the Azmaka femur tentatively to cf. Graecopithecus, partly on the basis of its age: the three specimens (the Greek mandible, a premolar from an earlier Azmaka horizon, and this femur) are all confined to roughly the first 70,000 years of the Messinian stage. They’re essentially contemporaries.
What actually makes this femur unusual is a cluster of features that, when you look at them individually, each tell a partial story, but together tell a stranger and more compelling one.
The femoral shaft is nearly straight. In African great apes the shaft curves anteroposteriorly, an adaptation that the team interprets as related to arboreal locomotion rather than terrestrial quadrupedalism. No actively moving terrestrial quadruped primate has a strongly curved femoral shaft. The Azmaka bone looks more like an australopith’s femur in this respect than like a chimpanzee’s. The greater trochanter sits slightly below the level of the femoral head, a configuration associated with increased hip mobility, also seen in some chimpanzees but more pronounced here in a direction consistent with early hominins.
The femoral neck is deep superoinferiorly, unusually so. Neck depth is associated with ground reaction forces at the hip, which is why human necks are deep too: walking upright loads the hip joint in a way that requires a structurally robust neck. The cortical bone distribution within the neck is asymmetrical, thick inferiorly and thin superiorly. This is the pattern you’d expect in an animal subject to consistent unidirectional loads on the hip, as in walking quadrupedally or bipedally. In suspensory apes like chimpanzees and orangutans, the cortical bone is distributed more evenly around the neck because forces come from multiple directions during climbing. The Azmaka specimen looks nothing like a suspensory ape in this respect. It looks like a biped, or at minimum like something doing a lot of stereotypical weight-bearing on its hind limbs.
Then there’s the feature the team calls FNOL: the femoral neck oblique length, which measures the ascending, superomedially oriented medial portion of the proximal neck margin. This is a geometric detail that sounds arcane until you understand what it represents. In hominins, the proximal margin of the femoral neck is divided into a short, nearly horizontal lateral part and a long, steeply ascending medial part. This elongated medial portion is what separates the femoral head from the greater trochanter and reflects the elongation of the neck itself, which biomechanically increases the moment arm of the hip abductor muscles. That’s important for stabilizing the pelvis when you’re standing on one leg, which is exactly what you do during every stride of bipedal walking. In non-hominin hominoids, this medial portion is shorter and usually slightly concave. In the Azmaka femur, it is long and straight, grouping statistically with Orrorin, Australopithecus afarensis, Paranthropus, and Homo sapiens, and not with any of the European Miocene apes.
Applying a discriminant function developed by Lovejoy and colleagues to distinguish bipedal hominids from knuckle-walking great apes, the Azmaka femur scores on the biped side of the boundary. Just barely, but clearly.
A gluteal tuberosity is present on the posterior shaft surface, vertical and posterolaterally placed. In African great apes, the gluteal tuberosity is effectively replaced by a lateral spiral pilaster, a different structure associated with a different configuration of the gluteus maximus, positioned more for hip extension and climbing rather than hip stabilization during upright posture. The Azmaka femur has no lateral spiral pilaster. Its gluteal configuration most closely resembles early hominins like Orrorin and Australopithecus afarensis. A faint intertrochanteric line is also present, associated with a well-developed iliofemoral ligament, which limits hip hyperextension and is characteristic of creatures that habitually extend their hips toward or past neutral.
None of these features, alone, is conclusive. Some appear in unexpected places. Gluteal tuberosities of varying development occur across many primates. A straight shaft is not unique to hominins. But the combination is coherent. The pattern points consistently in the same direction.
Not a modern walker, but something new
The team is explicit that this is not obligate bipedalism. A number of features associated with fully committed bipedalism in modern humans are absent or underdeveloped: there’s no hypotrochanteric fossa, no clear obturator externus groove, and the shaft is not platymeric (flattened front-to-back) in the way typical of later bipeds. The neck-shaft angle is about 122 degrees, which is in the range of australopiths and modern humans but lower than in the African great apes, lower than in Danuvius guggenmosi (a proposed Miocene biped from Germany with a 134-degree angle), and notably lower than the estimated values for Sahelanthropus, which apparently had an angle somewhere between 138 and 148 degrees, more consistent with orangutans and the suspensory European ape Hispanopithecus than with hominins.
The most accurate characterization the team offers is facultative bipedalism alongside terrestrial quadrupedalism, without the specialized arboreal features of extant great apes. This creature could walk upright. It probably also moved quadrupedally on the ground. It was not a habitual tree-swinger. The locomotor repertoire was complex, transitional, and unlike anything living today.
That transitional quality is important. The pattern of bipedal features in the Azmaka femur resembles early hominins more than it resembles later ones, a relationship the authors note recalls the distinction between australopiths and Homo. Bipedalism, in other words, appears to have been a multi-phase process. The Azmaka hominine may represent phase one.
Professor Spassov has described the bone’s suite of features as combining the characteristics of terrestrial quadrupeds and bipeds, clustering mostly with early bipeds but partially with African apes. Begun characterizes the specimen as a stage between arboreal ancestors like Danuvius and the later East African bipeds.
The comparison with Sahelanthropus is worth dwelling on. Sahelanthropus is currently the oldest proposed hominin at about 7 million years, slightly younger than the Azmaka specimen, from Chad. Daver and colleagues argued in 2022 that several features of the Sahelanthropus femur support habitual bipedalism. Cazenave and colleagues argued the opposite in 2025. The Azmaka team has looked carefully at the Sahelanthropus femur and reached a pointed conclusion: the high neck-shaft angle, the strongly curved shaft, the highly sigmoid shape of the lateral linea aspera lip, and extensive rodent gnawing that has obscured potentially diagnostic posterior surface features, all make the bipedal argument for Sahelanthropus substantially weaker than for the Azmaka specimen. The Azmaka hominine, the authors conclude, was probably more committed to bipedal behavior than Sahelanthropus, despite being slightly older.
Whether Sahelanthropus was bipedal remains actively contested and will not be settled here.
The question of where
Assuming the Azmaka femur is what the team says it is, an early biped older than Orrorinand contemporaneous with or older than Sahelanthropus, the next question is almost unsettling: what does it mean for a potential human ancestor to be in Bulgaria at 7.2 million years ago?
The Balkans and Anatolia were busy with hominines in the Late Miocene. Ouranopithecus macedoniensis from Greece, dated to around 9 to 9.6 million years ago. Anadoluvius turkaefrom central Turkey at about 8.7 million years. And now Graecopithecus from Greece and Bulgaria at 7.2 million years. This is not a random scattering. The Balkan-Iranian palaeozoogeographic province sits along the dispersal corridor between Eurasia and Africa, and during the Messinian it was experiencing dramatic environmental upheaval as the Mediterranean periodically restricted and hyperarid conditions developed across western Asia between roughly 7 and 6 million years ago.
Böhme and colleagues published an analysis in 2021 showing that mammalian dispersal between Eurasia and Africa in the Messinian moved almost exclusively from Eurasia to Africa, primarily via the Arabian Peninsula. The mammal faunas of Azmaka include a high diversity of savannah-adapted taxa consistent with open and bush savannah environments. The fauna at Pyrgos Vasilissis, the Greek locality for Graecopithecus, tells a similar ecological story. These were not forest apes. They were living in and adapted to open wooded landscapes.
The hypothesis the team advances is straightforward, if radical in its implications: the ancestors of Graecopithecus were Balkan-Anatolian hominines already adapted to semi-terrestrial life in open habitats. As those habitats became increasingly arid and patchy, some lineages dispersed southward into Africa, where they encountered and eventually occupied more forested settings as the ancestors of African apes and hominins. What we identify later as the earliest African hominins, including Orrorin and eventually Australopithecus, may represent descendants of this Eurasian population arriving in Africa rather than a purely African radiation.
This is a hypothesis, not a settled conclusion. The sample from Azmaka is small. Attribution to cf. Graecopithecus is parsimonious but not proven. One premolar, one mandible, one femur. The team cannot exclude the possibility that multiple hominine taxa coexisted in the earliest Messinian Balkans. A recently published molar from Veles in North Macedonia, dated to approximately the same interval, suggests at minimum that hominine presence in the Balkans during this period was more widespread than a single site would indicate.
The savannah environment, the open landscape, the terrestrial locomotion: these are consistent with the scenario in which open country conditions selected for a more diverse positional repertoire that included regular bipedalism, perhaps for foraging on the ground as arboreal resources became scarce, perhaps for carrying offspring or resources across gaps between tree patches, perhaps for the elevated vantage that upright posture provides in open terrain.
We do not know exactly what this animal looked like moving through a Late Miocene Bulgarian savannah. We have one femur, dated with good palaeomagnetic and biochronological constraints, from a female who weighed about as much as a large dog. The bone is dark, stained with manganese, encrusted with carbonate, cracked in places. In cross-section through the neck it looks startlingly like a small human femur, not like a chimpanzee’s.
That resemblance is what the argument turns on. And it is, in the judgment of a team that has looked at this specimen and the comparative anatomy carefully over several years, not coincidental.
Further Reading
Cazenave, M., Pina, M., Hammond, A. S., Böhme, M., Begun, D. R., Spassov, N., et al. (2025). Postcranial evidence does not support habitual bipedalism in Sahelanthropus tchadensis: A reply to Daver et al. (2022). Journal of Human Evolution, 198, 103557. https://doi.org/10.1016/j.jhevol.2024.103557
Daver, G., Guy, F., Mackaye, H. T., Likius, A., Boisserie, J. R., Moussa, A., et al. (2022). Postcranial evidence of late Miocene hominin bipedalism in Chad. Nature, 609, 94–100. https://doi.org/10.1038/s41586-022-04901-z
Fuss, J., Spassov, N., Begun, D., & Böhme, M. (2017). Potential hominin affinities of Graecopithecus from the Late Miocene of Europe. PLOS ONE, 12, e0177127.
Böhme, M., Spassov, N., Fuss, J., Tröscher, A., Deane, A. S., Prieto, J., et al. (2019). A new Miocene ape and locomotion in the ancestor of great apes and humans. Nature, 575, 489–493. [This is the Danuvius paper]
Böhme, M., Spassov, N., Majidifard, M. R., et al. (2021). Neogene hyperaridity in Arabia drove the directions of mammalian dispersal between Africa and Eurasia. Communications Earth & Environment, 2, 85. https://doi.org/10.1038/s43247-021-00158-y
Böhme, M., Spassov, N., Ebner, M., Geraads, D., Hristova, L., et al. (2017). Messinian Age and savanna environment of the possible hominin Graecopithecus from SE Europe. PLOS ONE, 12, e0177347.
Richmond, B., & Jungers, W. (2008). Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism. Science, 319, 1662. https://doi.org/10.1126/science.1154197
Lovejoy, C. O., Meindl, R., Ohman, J., Heiple, K. D., & White, T. (2002). The Maka Femur and Its Bearing on the Antiquity of Human Walking: Applying Contemporary Concepts of Morphogenesis to the Human Fossil Record. American Journal of Physical Anthropology, 119, 97–133.
Macchiarelli, R., Bergeret-Medina, A., Marchi, D., & Wood, B. (2020). Nature and relationships of Sahelanthropus tchadensis. Journal of Human Evolution, 149, 102898. https://doi.org/10.1016/j.jhevol.2020
Sevim-Erol, A., Begun, D. R., Yavuz, A., et al. (2023). A new ape from Türkiye and the radiation of late Miocene hominines. Communications Biology, 6, 842. https://doi.org/10.1038/s42003-023-05210-5
Stamos, P., & Alemseged, Z. (2023). Hominin locomotion and evolution in the Late Miocene to Late Pliocene. Journal of Human Evolution, 178, 103332.
Almécija, S., Tallman, M., Alba, D. M., Pina, M., Moyà-Solà, S., & Jungers, W. L. (2013). The femur of Orrorin tugenensis exhibits morphometric affinities with both Miocene apes and later hominins. Nature Communications, 4, 2888. https://doi.org/10.1038/ncomms3888
Pina, M., Alba, D. M., Moyà-Solà, S., & Almécija, S. (2019). Femoral neck cortical bone distribution of dryopithecin apes and the evolution of hominid locomotion. Journal of Human Evolution, 136, 102651.
Spassov, N., Youlatos, D., Böhme, M., Bogdanova, R., Hristova, L., & Begun, D. R. (2026). An early form of terrestrial hominine bipedalism in the Late Miocene of Bulgaria. Palaeobiodiversity and Palaeoenvironments. https://doi.org/10.1007/s12549-025-00691-0







